Las membranas son las que generan los compartimientos celulares.
Básicamente RE, mitocondria (con doble membrana), núcleo (con doble sistema de membrana), peroxisomas, lisosomas y las vesículas emergentes del Golgi o las vesículas endocíticas.
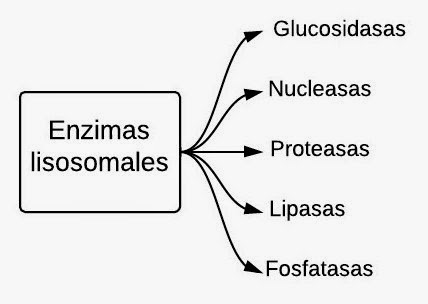
Lisosomas.
 |
| En esta imagen vemos un lisosoma que tiene adentro una mitocondria y un peroxisoma→es secundario porque tiene algo adentro. | | | |
La función del lisosoma es degradar cosas. Están llenos de enzimas hidrolíticas (en presencia de agua hidrolizan proteínas en aminoácidos, lípidos en ácidos grasos, glúcidos complejos en glúcidos simples). Estas enzimas hidrolíticas tienen la particularidad de que trabajan a un pH al que ninguna otra enzima trabaja, trabajan a pH ácido. El lisosoma tiene un pH interno 4-5. Para mantener este pH interno ácido, el lisosoma tiene en su membrana una
bomba de protones que permite el ingreso de protones desde el citosol hacia la luz del lisosoma. Además poseen
canales de cloruro, porque no pueden entrar solamente protones (deben mantener la electroneutralidad).
La membrana del lisosoma es la única que posee una bomba de protones, por lo tanto esta bomba de protones es un
marcador bioquímico. También posee una proteína: LAMP2. Las proteínas LAMP2 son proteínas asociadas a la membrana lisosomal, es una proteína típica de lisosomas, por lo tanto también es un marcador bioquímico.
 |
| Confocal immunofluorescent analysis of LAMP2 antibody with HepG2 cell. |
Ejemplo: cuando hay una mutación en una enzima que es una glucosidasa, una hexoaminidasa A, hay un glucolípido, que tiene una ceramida, que no se degrada. Cuando no se degrada se acumula dentro del lisosoma, y los lisosomas se empiezan a llenar de gangliósidos, y así empiezan a ser no funcionales. Así se genera una enfermedad neurodegenerativa que se denomina
enfermedad de Tay-Sachs.
Los lisosomas cuando se fusionan con lo que tienen que degradar forman lo que se llama lisosoma secundario. Se fusionan con moléculas solubles que son llevadas dentro de la célula por endocitosis, formando así una vesícula endocítica, que luego forma un endosoma temprano y un endosoma tardío. Cuando el lisosoma se fusiona con el endosoma, tenemos un lisosoma secundario. Pero también el lisosoma es capaz de degradar organelas o compartimientos viejos, y esto se llama
autofagosoma, y este proceso se denomina
autofagia. Los lisosomas también pueden fusionarse son fagosomas que contienen bacterias.
Peroxisomas.
Los peroxisomas, en lugar de tener enzimas hidrolíticas como tienen los lisosomas, tienen enzimas oxidativas: oxidan todo lo que pasa por ahí. En general oxidan ácidos grasos de cadenas muy largas, porque la mitocondria hace β-oxidación pero solo de cadenas de 20-24 C, no más. Pero hay ácidos grasos que son de cadenas muy largas y esto debe ser oxidado en el peroxisoma.
La membrana del peroxisoma tiene un transportador de tipo ABC que permite el transporte del ácido graso del citosol hacia el interior del peroxisoma y aquí el peroxisoma lo oxida a un ácido graso que tiene dos átomos de C menos, se genera un acetilo y un ácido graso más corto. Para esta oxidación se necesita oxígeno y como consecuencia de la oxidación se produce peróxido de hidrógeno (H
2O
2).
El H
2O
2 es tóxico para la célula, por lo tanto, rápidamente esta enzima catalaza va a degradar el H
2O
2 en H
2O y O
2.
La catalasa es un
marcador bioquímico. Con la catalasa no se puede identificar a la membrana del peroxisoma, se puede identificar al peroxisoma.
Cuando el transportador de la membrana del peroxisoma que se encarga del transporte del ácido graso al interior del peroxisoma no funciona, el ácido graso no entra, por lo tanto no puede ser degradado, pero el problema es que se acumula en la célula y se produce una enfermedad que se conoce como
adrenoleucodistrofia (ADL), la famosa enfermedad de Lorenzo. Es una enfermedad ligada al cromosoma X (es transmitida por las mujeres).
RE y aparato de Golgi.
 |
| Microfotografía electrónica donde se ve un RE muy desarrollado y todas las vesículas secretorias. |
 |
| Acá se ve el retículo endoplasmático marcado con una proteína específica del RE que es el PDI (proteína disulfuro isomerasa), que es la encargada de poner los puentes disulfuro cuando se pliegan las proteínas. |
Mitocondrias.
Doble membrana:
Funciones:
•
Producción de ATP → función más importante
• β-oxidación de ácidos grasos
• Ciclo de la urea (degradación de ácidos grasos)
• Depósito de Ca
2+ → es el principal depósito de Ca
2+ de las células. Este depósito de Ca
2+ no es fácilmente movilizable, el Ca
2+ que se moviliza es es del RE.
Núcleo.
El núcleo es el gran compartimiento que contiene al genoma. Está rodeado por un doble sistema de membranas. Aquí ocurre la transcripción y maduración del ARNm, ARNt y el ARNr, se ensamblan los ribosomas. Dentro del núcleo ocurren también procesos de biosíntesis y biodegradación. La síntesis del ARN ocurre dentro del núcleo, pero también hay síntesis de lípidos.
El núcleo tiene un sistema del tipo proteasómico para la degradación.
Citoesqueleto.
Los compartimientos vienen dados por las membranas pero la forma de la célula viene dada por el citoesqueleto.
 |
| Enterocito | |
 |
| Fibroblasto |